Aluminio en el tejido cerebral de sujetos autistas.

Revista de oligoelementos en medicina y biología
Autores: Matthew Mould, Dorcas Umar, Andrew King, Christopher Exley
26 November 2017
Resumen
El trastorno del espectro autista es un trastorno del desarrollo neurológico de etiología desconocida (causa). Se sugiere que la causa involucra tanto la susceptibilidad genética como los factores ambientales, que también incluyen toxinas ambientales al final. Se han realizado intentos para vincular la exposición humana a la toxina ambiental de aluminio con el trastorno del espectro autista. Aquí hemos usado la técnica de espectrometría de absorción atómica de horno de grafito calentado transversalmente (espectrometría de absorción atómica de un horno de grafito calentado transversalmente) para medir, por primera vez, el contenido de aluminio del tejido cerebral de sujetos donantes con diagnóstico de autismo. También utilizamos flúor selectivo para aluminio para identificar aluminio en el tejido cerebral mediante microscopía de fluorescencia. El contenido de aluminio del tejido cerebral en el autismo ha sido decididamente alto. La media (entre paréntesis, la desviación estándar) del contenido de aluminio en los 5 individuos para cada lóbulo fue 3,82 (5,42), 2,30 (2,00), 2,79 (4,05) y 3,82, 5,17 (15) μg / g de peso de materia seca para los lóbulos occipital, frontal, temporal y parietal, respectivamente. Estos se encuentran entre los valores más altos de aluminio en el tejido cerebral humano medidos hasta ahora y uno tiene que preguntarse por qué, por ejemplo, el contenido de aluminio del lóbulo occipital de un niño de 8,74 años debe ser de 11,59 (10) μg / g de peso de materia seca? Se usó microscopía de fluorescencia selectiva de aluminio para identificar aluminio en el tejido cerebral de XNUMX donantes. Si bien se imaginó que el aluminio estaba asociado con las neuronas, se observó que está presente en la zona intracelular en células similares a la microglia y en otras células inflamatorias no neuronales de las meninges, vascularización, materia gris y sustancia blanca. La preeminencia del aluminio intracelular asociado con células no neuronales ha sido una observación excepcional en el tejido cerebral de sujetos autistas y puede ofrecer pistas tanto sobre el origen del aluminio presente en el cerebro como sobre el posible papel de esta sustancia en causar el trastorno del espectro autista
Introducción
El trastorno del espectro autista (TEA) es un grupo de afecciones patológicas del desarrollo neurológico de causa desconocida. Es muy probable que los factores genéticos [1] y ambientales [2] estén asociados con el inicio y el progreso de ASD, mientras que se espera que los mecanismos subyacentes a su ASI sean multifactoriales [3-6]. Se sospecha que la exposición humana al aluminio es la causa del TEA, pero las conclusiones han sido cuestionables [7-10]. Hasta ahora, la mayoría de los estudios han utilizado el cabello como un indicador de exposición humana al aluminio, mientras que el contenido de aluminio en la sangre y la orina se ha utilizado en un grado mucho más limitado. Las vacunas pediátricas que incluyen un adyuvante a base de aluminio son una medida indirecta de la exposición de los lactantes al aluminio y su uso cada vez mayor se ha relacionado directamente con la mayor prevalencia de TEA [11]. Los modelos animales de ASD continúan apoyando una conexión con aluminio y adyuvantes a base de aluminio utilizados particularmente para vacunas humanas [12]. Hasta la fecha, no hay estudios sobre la presencia de aluminio en el tejido cerebral de los donantes que murieron con un diagnóstico de TEA. Medimos la cantidad de aluminio presente en el tejido cerebral de sujetos autistas e identificamos la ubicación del aluminio en estos tejidos.
Materiales y métodos
2.1. Medición de las cantidades de aluminio en los tejidos cerebrales.
Se obtuvo la aprobación ética junto con los tejidos del "Oxford Brain Bank" (Oxford Brain Bank - 15 / SC / 0639). Se obtuvieron muestras de corteza igual a aproximadamente un gramo de materia congelada de los lóbulos temporal, frontal, parietal y occipital y del hipocampo (solo 0,3 gr) de 5 individuos con diagnóstico de ASD confirmado por ADI-Rconfirmed (Autism Diagnostic Interview -Revisado), 4 hombres y 1 mujer, de 15 a 50 años (Tabla 1). El contenido de aluminio de estas telas se midió por medio de un método consolidado y totalmente validado [13] que solo se describe brevemente aquí.
Resultados
3.1 Contenido de aluminio en los tejidos cerebrales.
El contenido de aluminio en todos los tejidos osciló entre 0,01 (el límite de cuantificación) y 22,11 mg / g de peso de materia seca. (Tabla 1). El contenido de aluminio para el cerebro en su conjunto (n = 4 o 5 dependiendo de la disponibilidad de tejido del hipocampo) osciló entre 1,20 (1,06) mg / g de materia seca para la mujer de 44 años (A1 ) a 4,77 (4,79) mg / g del varón de 33 años (A5). Las mediciones previas de aluminio en el cerebro, incluido nuestro estudio de 60 cerebros [15], nos permitieron definir aproximadamente las clases de contenido de aluminio comenzando con valores menores o iguales a 1,00 mg / g como patológicamente benignos (al contrario del concepto de 'normal'). Aproximadamente el 40% de los tejidos (24/59) tenían un contenido de aluminio considerado patológicamente preocupante (³2,00 mg /) mientras que alrededor del 67% de estos tejidos tenían un contenido de aluminio considerado patológicamente significativo (³3,00 mg / g ). Los cerebros de los 5 individuos tienen al menos un tejido con un contenido de aluminio patológicamente significativo. Los cerebros de los 4 individuos tenían al menos un tejido con un contenido de aluminio mayor o igual a 5,00 mg / g, mientras que 3 de estos tenían al menos un tejido con un contenido de aluminio mayor o igual a 10,00 mg / g (Cuadro 1). El contenido medio de aluminio (desviación estándar entre paréntesis) de los 5 individuos para cada lóbulo fue 3,82 (5,42), 2,30 (2,00), 2,79 (4,05) y 3,82 ( 5,17) para los lóbulos frontal, occipital, frontal, temporal y parietal. No hubo diferencias estadísticamente significativas en el contenido de aluminio entre los 4 lóbulos.
3.2. Fluorescencia debida al aluminio en los tejidos cerebrales.
Examinamos secciones en serie del cerebro de 10 individuos (3 mujeres y 7 hombres) que murieron con un diagnóstico de TEA y registraron la presencia de aluminio en estos tejidos (Tabla S1). La excitación del complejo de aluminio y lumogallion emite una fluorescencia naranja característica que aparece cada vez más de color amarillo brillante a medida que aumenta la intensidad de la fluorescencia. El aluminio, identificado como depósitos reactivos de lumogallion, se registró en al menos un tejido en los 10 individuos. La autofluorescencia de las secciones en serie inmediatamente adyacentes confirmó que la fluorescencia de lumogallion es indicativa de la presencia de aluminio. Los depósitos de aluminio fueron significativamente más frecuentes en hombres (129 de 7 individuos) que en mujeres (21 de 3 individuos). Se encontró aluminio tanto en la sustancia blanca (62 depósitos) como en la materia gris (88 depósitos). En las mujeres, la mayoría de los depósitos de aluminio se identificaron como extracelulares (15 de 21), mientras que en los hombres se encontró la situación opuesta con 80 de 129 depósitos en el área intracelular. Solo se nos proporcionaron 3 secciones en serie de cada tejido y, por lo tanto, no pudimos realizar ninguna coloración para identificar la morfología general, lo que significa que no siempre fue posible determinar qué subtipo de células mostraba fluorescencia debido al aluminio. . Se identificaron glóbulos blancos mononucleares cargados con aluminio, probablemente linfocitos, en las meninges y probablemente ingresaron al tejido cerebral desde el sistema linfático (Fig. 10). El aluminio se podía ver claramente dentro de las células o en forma de depósitos puntuales discretos, o como fluorescencias amarillas intensas (Fig. 1). El aluminio se ha localizado en las células inflamatorias asociadas con la vascularización (Fig. 2). En un caso, se observó lo que parece ser un linfocito o monocito cargado de aluminio dentro de un vaso sanguíneo rodeado de glóbulos rojos, mientras que otro linfocito probable que muestra fluorescencia amarilla intensa se observó en la membrana adventicia (Fig. 2b). Las células gliales, incluidas algunas similares a las de microglia que mostraron fluorescencia debido a la presencia de aluminio, a menudo se han observado en el tejido cerebral cerca de depósitos extracelulares coloreados por aluminio (Figuras 3 y 4). Los depósitos de aluminio de aproximadamente 1 mm de diámetro eran claramente visibles tanto en el cuerpo redondo como en el ameboide de las células gliales (p. Ej. Fig. 3b). El aluminio intracelular se ha identificado en neuronas y células probables similares a las de la glía, y a menudo en las proximidades o en el mismo sitio que la lipofuscina (Fig. 5). La fluorescencia selectiva para aluminio trabajó en la identificación de aluminio en sitios extracelulares e intracelulares en células neuronales y no neuronales y en todos los tejidos cerebrales estudiados (Figuras 1-5). El método identifica el aluminio solo como lo demuestran grandes áreas de tejido cerebral sin ninguna fluorescencia característica que indique la positividad al aluminio (Fig.
Hilo
El contenido de aluminio del tejido cerebral de donantes diagnosticados con TEA fue extremadamente alto (Tabla 1). Si bien se encontró una variabilidad significativa entre diferentes tejidos, diferentes lóbulos y diferentes sujetos, el contenido promedio de aluminio para cada lóbulo (entre 5 individuos) estuvo en los niveles más altos de todas las mediciones previas del contenido de aluminio en el cerebro, incluidos los casos de trastornos iatrogénicos como la encefalopatía por diálisis [13,15, 16-19]. Los 4 donantes masculinos tenían concentraciones de aluminio en el cerebro más altas que el único donante femenino. En estos machos autistas, registramos algunos de los valores de contenido de aluminio más altos en el cerebro jamás medidos en tejidos sanos o enfermos, incluidos valores de 17,10 18,57 y 22,11 mg / g de materia seca. (Tabla 1). Lo que distingue estos datos de otros análisis del aluminio en el cerebro en otras enfermedades es la edad de los individuos autistas. ¿Por qué, por ejemplo, un niño de 15 años tendría un contenido de aluminio tan alto en el tejido cerebral? No hay datos comparables en la literatura científica, siendo los datos igualmente altos más cercanos el de un hombre de 42 años con una "forma familiar" de la enfermedad de Alzheimer (FAD) [19]. La microscopía de fluorescencia selectiva de aluminio proporcionó indicaciones de sitios de deposición de aluminio en estos tejidos cerebrales de sujetos autistas (Figuras 1-5).
El aluminio se ha encontrado tanto en la materia blanca como en la gris y en los sitios extracelulares y celulares. Estos últimos estaban particularmente extendidos en estos tejidos de sujetos autistas. Las células que morfológicamente parecían no neuronales y muy cargadas con aluminio se han identificado como células asociadas con las meninges (Fig. 1), vascularización (Fig. 2) y como células de la materia gris y la sustancia blanca (Figuras 3-5). Algunas de estas células parecían gliales (probablemente astrocitos), mientras que otras tenían núcleos alargados que les daban la apariencia de células microglia [5]. Estos últimos a veces se han visto cerca de depósitos extracelulares de aluminio. Esto implica que el aluminio de alguna manera cruzó la barrera hematoencefálica y fue tomado de una célula nativa, una célula de microglia. Curiosamente, la presencia ocasional de células inflamatorias cargadas de aluminio en la vascularización y leptomeninges abre la posibilidad de una forma diferente de entrada de aluminio en el cerebro, o intracelularmente. Sin embargo, para que este segundo escenario sea válido, uno esperaría que ocurriera algún tipo de daño intracerebral para permitir el escape de linfocitos y monocitos de la vascularización. La identificación realizada aquí de las células no neuronales, incluidas las células inflamatorias, las células gliales y las células de la microglia, todas cargadas con aluminio, es una observación excepcional para TEA. Por ejemplo, la mayoría de los depósitos de aluminio identificados en el tejido cerebral en las "formas familiares" de la enfermedad de Alzheimer fueron extracelulares y casi siempre se asociaron con materia gris [19].
El aluminio es citotóxico [21] y su asociación (mostrada aquí) con las células inflamatorias de la vascularización, las meninges y el sistema nervioso central difícilmente puede ser benigna. La microglia de aluminio muy cargada, si bien puede seguir siendo viable, al menos durante un tiempo, inevitablemente se verá comprometida y se cree que la disfunción de la microglía está involucrada en la etiología de los TEA [22], por ejemplo, en el daño de la falta de hojas sináptica [23] . Además, el hecho de que estos datos sugieran que la entrada de aluminio en el cerebro por las células del sistema inmunitario que circulan en la sangre y la linfa se acelera en los sujetos autistas podría comenzar a explicar la pregunta planteada primero sobre por qué Hay tanto aluminio en el cerebro de un niño autista de 15 años. Una limitación de nuestro estudio es el pequeño número de casos disponibles para el análisis y la disponibilidad limitada de tejido. Con respecto al último factor, tener acceso a solo 1 g de tejido congelado y solo 3 secciones en serie de tejido fijado por lóbulo normalmente podría considerarse una limitación significativa. Ciertamente, si no hubiéramos identificado ningún depósito significativo de aluminio en una muestra de tejido tan pequeña (la masa promedio del cerebro varía entre 1.500 y 2.000 g), ese resultado sería equívoco. Sin embargo, el hecho de que hayamos encontrado aluminio en cada muestra de tejido, congelada o congelada, sugiere que las personas diagnosticadas con TEA tienen niveles extraordinariamente altos de aluminio en el tejido cerebral y que este aluminio está asociado de manera preeminente con células no neuronales incluidas los de microglia y otros monocitos inflamatorios.
Conclusiones
Hemos realizado la primera medición del contenido de aluminio en el tejido cerebral en sujetos autistas y hemos demostrado que el contenido de aluminio en el cerebro es extraordinariamente alto. Identificamos aluminio en el tejido cerebral en las áreas extracelular e intracelular (tanto en neuronas como en células no neuronales). La presencia de aluminio en las células inflamatorias en las meninges, en la vascularización, en la materia gris y blanca es una observación excepcional y podría demostrar un papel causante del aluminio en la etiología de los TEA.
PSUR 16: duplicación de las muertes esperadas
Si todos los niños que recibieron la primera dosis de la vacuna reciben un total de cuatro dosis y la última dosis se administra en el segundo año de vida, entonces se puede estimar que una cuarta parte (25%) de las dosis se administra a niños mayores a un año Este es el calendario de vacunación recomendado en Alemania. Sin embargo, algunos países, como Italia, recomiendan solo tres dosis, todas en el primer año y ninguna en el segundo. Además, no todos los niños reciben todas las dosis recomendadas. Por lo tanto, es poco probable que el 20-25% de las dosis se usen en el segundo año. En la PSUR 15, se estimó que el 90,6% de las dosis vendidas se usaron en lactantes menores de un año y el 9,4% en los mayores de un año. En PSUR 16, la estimación de las dosis recibidas en el segundo año se ha más que duplicado (del 9,4% al 20%) y, por lo tanto, la estimación de las muertes esperadas se ha duplicado. A pesar de la duplicación de las muertes esperadas, el número de muertes observadas en el segundo año fue mayor de lo esperado en los primeros 3 días después de la vacunación (Tabla 36, p249). Si la estimación en PSUR 15 de que el 9,4% de las dosis se usan en el segundo año es correcta, esto también se aplica a PSUR 16 y, por lo tanto, las muertes observadas son más altas que las esperadas en los primeros 7 días.
Conflicto de intereses de interés.
Los autores declaran que no tienen conflictos de intereses.
Gracias
La investigación está respaldada por fondos de Instituto de Investigación de Seguridad Médica Infantil (CMSRI), una fundación sin fines de lucro con sede en Washington DC, EE. UU. que se ocupa de la investigación
Referencias
- A. Krishnan, R. Zhang, V. Yao, CL Theesfeld, AK Wong et al., Predicción de todo el genoma y caracterización funcional de la base genética del trastorno del espectro autista, Nature Neuroscience 19 (2016) 1454-1462.
- LA Sealey, BW Hughes, AN Sriskanda, JR Guest, AD Gibson et al., Factores ambientales en el desarrollo de trastornos del espectro autista, Environ. Int.88 (2016) 288-298.
- R. Koyama, Y. Ikegaya, Microglia en la patogénesis de los trastornos del espectro autista, Neurosci. Res.100 (2015) 1-5.
- P. Li, JM. Zhou, El eje microbiota-intestino-cerebro y su posible papel terapéutico en el trastorno del espectro autista, Neuroscience 324 (2016) 131-139.
- C. Kaur, G. Rathnasamy, EA. Ling, Biología de microglia en el cerebro en desarrollo, J. Neuropathol Exp. Neurol. 76 (2017) 736-753.
- M. Varghese, N. Keshav, S. Jacot-Descombes, T. Warda, B. Wicinski et al., Trastorno del espectro autista: neuropatología y modelos animales, Acta Neuropathol. 134 (2017) 537-566.
- H. Yasuda, Y. Yasuda, T. Tsutsui, Estimación de niños autistas por análisis de metallomics, Sci. Rep. 3 (2013) 1199.
- FEB Mohamed, EA Zaky, AB El-Sayed, RM Elhossieny, SS Zahra et al., Evaluación del cabello de aluminio, plomo y mercurio en una muestra de niños egipcios autistas: factores de riesgo ambientales de metales pesados en el autismo, Neurol conductual. (2015) Art. 545674.
- MH Rahbar, M. Samms-Vaughn, MR Pitcher, J. Bressler, M. Hessabi et al., Papel de los genes metabólicos en las concentraciones de aluminio en sangre de niños jamaicanos con y sin trastorno del espectro autista, Int. J. Environ. Res. Public Health 13 (2016) 1095.
- AV Skalny, NV Simashkova, TP Klyushnik, AR Grabeklis, IV Radysh et al., Análisis de oligoelementos capilares en niños con trastornos del espectro autista y trastornos de la comunicación, Trace Elem. Med. Biol. 177 (2017) 215-223.
- L. Tomljenovic, CA Shaw, ¿Los adyuvantes de la vacuna de aluminio contribuyen a la creciente prevalencia del autismo ?, J. Inorg. Biochem. 105 (2011) 1489-1499.
- CA Shaw, Y. Li, L. Tomljenovic, la administración de aluminio a ratones neonatales en cantidades relevantes para la vacuna se asocia con resultados neurológicos adversos a largo plazo, J. Inorg. Biochem. 128 (2013) 237-244
- E. House, M. Esiri, G. Forster, P. Ince, C. Exley, Aluminio, hierro y cobre en tejidos del cerebro humano donados al estudio de función cognitiva y envejecimiento del consejo médico, Metallomics 4 (2012) 56-65.
- M. Mold, H. Eriksson, P. Siesjö, A. Darabi, E. Shardlow, C. Exley, Identificación inequívoca de adyuvante intracelular de aluminio en una línea celular monocítica THP-1, Sci. Rep. 4 (2014) 6287.
- A. Mirza, A. King, C. Troakes, C. Exley, La identificación de aluminio en el tejido cerebral humano utilizando microscopía de lumogallion y fluorescencia, J. Alzh. Dis. 54 (2016) 1333-1338.
- C. Exley, M. Esiri, Angiopatía congofílica cerebral severa coincidente con aumento de aluminio cerebral en un residente de Camelford, Cornwall, Reino Unido, J. Neurol. Neurosurg. Psiquiatría 77 (2006) 877-879.
- C. Exley, ER House, Aluminio en el cerebro humano, Monatsh. Chem. 142 (2011) 357-363.
- C. Exley, T. Vickers, Aluminio cerebral elevado y enfermedad de Alzheimer de aparición temprana en un individuo expuesto ocupacionalmente al aluminio: informe de un caso, J. Med. Case Rep. 8 (2014) 41.
- A. Mirza, A. King, C. Troakes, C. Exley, aluminio en el tejido cerebral en la enfermedad de Alzheimer familiar, J. Trace Elem. Med. Biol. 40 (2017) 30-36.
- R. Shechter, O. Miller, G. Yovel, N. Rosenzweig, A. London et al., El reclutamiento de macrófagos M2 beneficiosos para la médula espinal lesionada es orquestado por el plexo coroideo cerebral remoto, Immunity 38 (2013) 555-569.
- C. Exley, La toxicidad del aluminio en humanos, Morphologie 100 (2016) 51-55.
- MW Salter, B. Stevens, Microglia emerge como jugadores centrales en la enfermedad cerebral, Nat. Med.23 (2017) 1018-1027.
- U. Neniskyte, CT Gross, Jardineros errantes: poda sináptica dependiente de células gliales y trastornos del neurodesarrollo, Nat. Rev. Neurosci. 18 (2017) 658-670.
- Un enfoque alternativo a las vacunas combinadas: administración intradérmica de componentes aislados para el control de ántrax, botulismo, peste y shock tóxico estafilocócico (publicado en Revista de vacunas y terapias inmunitarias. 2008 3 de septiembre; 6: 5; autores Morefield GL, Tammariello RF y otros; https://www.ncbi.nlm.nih.gov/pubmed/18768085/)
Figura 1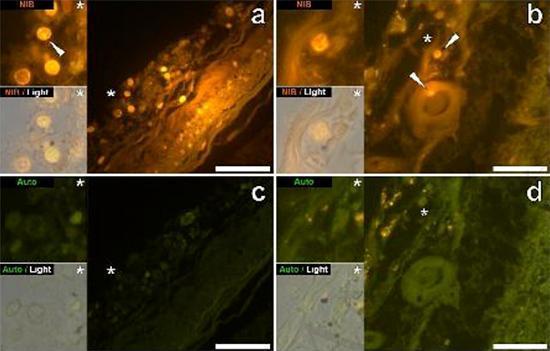
Figura 2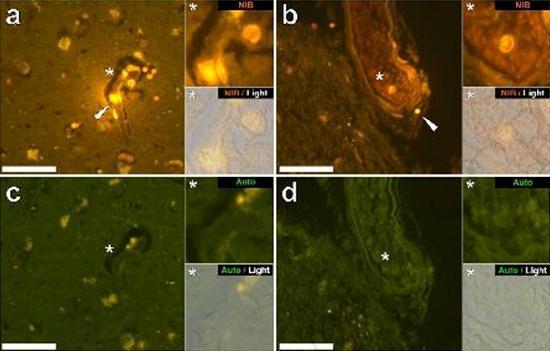
Figura 3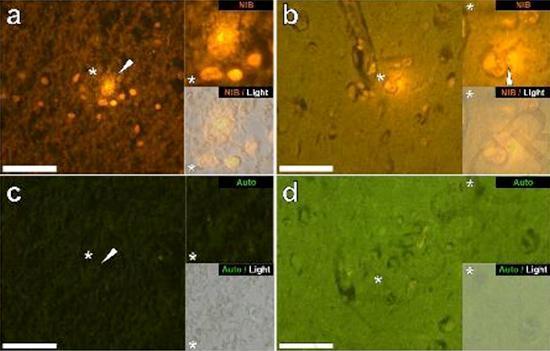
Figura 4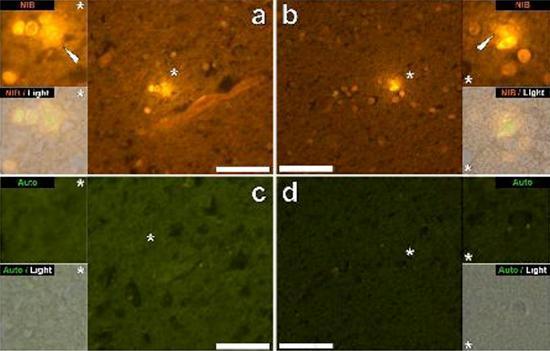
Figura 5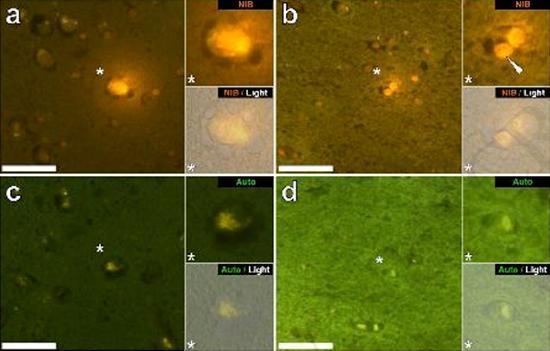
fuente: Aluminio en tejido cerebral en autismo
Descargar: Aluminio en tejido cerebral en autismo